
Comparative Single-Cell Transcriptome Analysis of Early Innate Lymphoid Cell Development in Humans and Mice
| Received 08 Oct, 2024 |
Accepted 29 Jan, 2025 |
Published 30 Jan, 2025 |
Background and Objective: Innate lymphoid cells (ILCs) are key immune cells involved in immune balance, pathogen defense, and tissue repair. However, their development, particularly in humans, is not well understood. This study maps ILC development in the mouse fetal liver and compares it with human ILC development, focusing on species-specific differences. Materials and Methods: This study compares ILC development between humans and mice to identify species-specific differences. Single-cell RNA sequencing data from human fetal tissues were obtained from GEO (GSE163587), while adult mouse bone marrow data (GSE113767) were used to identify ILC progenitors, including ALP, sEILP, cEILP, ILCP, and ILC2P. Homologous gene pairs between species were found using the bioMart package in R. Canonical correlation analysis (CCA) integrated the datasets, followed by PCA and UMAP for dimensionality reduction. Differential gene expression and functional enrichment were analyzed with DESeq2 and cluster Profiler. Pseudotime analysis using Monocle2 was used to infer the developmental trajectories of ILC populations in both species. Results: Current findings indicate the presence of an ILC developmental pathway in the mouse fetal liver that shares similarities with that in the bone marrow, demonstrating even more pronounced cellular proliferation in the fetal liver. The comparison between humans and mice revealed both conserved and divergent aspects of ILC development at cellular and molecular levels, including the regulation of key transcription factors and specific molecular expression patterns. Notably, this study uncovered distinct ILC subtypes specific to the embryonic stage in humans, which are postulated to play significant roles in the immune ecology of various tissues and organs. Conclusion: These discoveries enhance our understanding of ILC developmental processes and provide valuable theoretical support and practical guidance for research on the in vitro regeneration and application of ILCs.
INTRODUCTION
Innate lymphoid cells (ILCs) are a type of lymphocyte that lacks the expression of specific receptors for T-cell and B-cell surface antigens1. They primarily reside in mucosal tissues such as the intestine, lungs, and skin. These cells not only play a crucial role in clearing pathogen infections and maintaining immune homeostasis, but also contribute significantly to the formation of lymphoid organs, tissue repair, and immune-related metabolic diseases2,3. Based on the transcription factors they rely on for differentiation and the cytokines they secrete, ILCs are mainly classified into three groups: Group 1 ILCs (including natural killer cells and ILC1), Group 2 ILCs (ILC2), and Group 3 ILCs (ILC3 and lymphoid tissue-inducer cells). In adult mice, ILC development begins with common lymphoid progenitors in the bone marrow, progresses through an early innate lymphoid progenitor stage, and ultimately specializes in the innate lymphoid cell progenitor (ILCP) for ILC fate specification4-6. However, the embryonic origin and developmental pathway of mouse ILCs remain poorly understood. In recent years, the application of single-cell technology has provided new insights into the origin and hierarchy of early human ILC development. Embryonic ILCs originate from IL3RA+ lymphoid progenitors in the fetal liver and then specialize into ILCPs. Nevertheless, the similarities and differences in early ILC development between humans and mice are still unclear. Clarifying these species-specific differences and revealing the regulation of human-specific ILC development is not only scientifically valuable for understanding ILC development but also significant for guiding the in vitro regeneration of ILCs6,7.
To address these scientific questions, This study further explored published data on mouse fetal livers and constructed a developmental map of ILCs in mouse fetal livers for the first time. This study ILC-related populations in the fetal liver (including ALP, sEILP1, sEILP2, cEILP, and ILCP) that are similar to those in the bone marrow, as well as comparable fate specialization pathways8. One pathway leads to ILC specialization (ALP-sEILP1-cEILP-ILCP), while the other leads to dendritic cell fate specialization (ALP>sEILP1>sEILP2). However, there are differences between the fetal liver and bone marrow. Compared to the bone marrow, lymphoid progenitor populations and ILCP precursor cells in the fetal liver are in a more proliferative cell cycle state.
The study further compared the species-specific differences in ILC development between humans and mice. This study analyzed the specialization pathway of human MLP1>MLP2>ILCP/MLP3 for ILCs or dendritic cells. Integration and comparative analysis with single-cell sequencing data on early human ILC development, revealed the corresponding relationships and differences between human and mouse ILC developmental populations9,10. Specifically, human MLP1 corresponds to mouse ALP and sEILP1, both expressing characteristic genes of lymphoid progenitors; human MLP3 corresponds to mouse sEILP2; and human ILCP corresponds to mouse ILCP. Interestingly, human MLP2 and mouse cEILP are upstream of human and mouse ILCPs, respectively, but they do not have a good correspondence across species11-14. These results demonstrate the conservation and divergence of developmental hierarchies in human and mouse ILCs. Further compares the molecular expression patterns during ILC specialization in humans and mice. During ILC specialization, myeloid differentiation-related transcription factors such as Lmo2, Spi1, and Lyl1 are downregulated in both species, while key transcription factors for ILC development, such as Id2, Tox2, Tcf7, and Gata3, are upregulated. This suggests conserved transcriptional regulation during human and mouse ILC specialization. However, there are also species-specific differences in ILC development. For example, Bcl11a and Tcf4 expression gradually decreases in mice but remains unchanged in humans. Additionally, Zbtb16 and Rora are upregulated during mouse ILC specialization but not in humans, indicating differences in molecular regulation during early ILC development between the two species15.
Finally, the study focused on the tissue and organ specialization of human ILCs and analyzed their spatiotemporal heterogeneity. This study found that embryonic ILCs have a more active cell cycle and higher transcriptional activity compared to adult ILCs. Furthermore, the presence of embryonic-specific ILC subtypes with tissue and organ preferences among the three ILC subtypes was discovered. Pseudo-temporal analysis also revealed a fate specialization branch in all three major ILC types. One branch leads to ILC maturation, while the other specializes in embryonic-specific ILC subtypes. These findings suggest the existence of embryonic-specific tissue and organ ILC subtypes that may play crucial roles in the development of specific tissues and organs and their immune ecology.
MATERIALS AND METHODS
Study area and duration: The study was conducted at the First People's Hospital of Linping District, located in Hangzhou, China. The research spanned a period from September, 2021 to June, 2024.
Data collection: In their study, cells were isolated from human fetal hematopoietic (liver), lymphoid (thymus and spleen), and non-lymphoid (intestine, skin, and lung) tissues at 8, 10, and 12 post-conception weeks (PCW) using a single-cell RNA sequencing (scRNA-seq) strategy. They constructed a spatiotemporal and hierarchical developmental map of innate lymphoid cells in human embryos. The data is sourced from the Gene Expression Omnibus (GEO) under the accession number GSE163587, and it can be downloaded from the following link: GSE163587. Hence, sequencing analysis on adult mouse bone marrow using 10X transcriptome sequencing technology was conducted. This analysis provided data on the innate lymphoid cell progenitors present in adult mouse bone marrow, including populations such as ALP, sEILP, cEILP, ILCP, and ILC2P. The data is available from GEO under the accession number GSE113767 and can be downloaded from the following link: https://www,ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE113767.
Homologous gene conversion: The bioMart package is an R language interface that connects to the biomart database, allowing users to access it freely. This package enables various gene conversions. This studyutilized the biomart function from the Biomart software to process the gene names in the expression matrix. Using the use Mart function. This study downloaded gene sets for both humans and mice, followed by the use of the get LDS function to generate a comparative reference set of homologous gene pairs between humans and mice. Subsequently, the mouse gene names in the dataset to their corresponding human gene names were converted.
Batch effect integration: To integrate the data, canonical correlation analysis (CCA) was employed. Firstly, used the Find Integration Anchors function to identify integration anchors with the default parameters. Then, utilized the Integrate Data function to combine the anchored data, setting the parameters to “dims=1:30, k. weight=50.” Subsequently, normalized the data in the integrated space and performed Principal Component Analysis (PCA). The first 20 principal component dimensions were then subjected to UMAP dimensionality reduction, with the parameters set to “reduction='pca', dims=1:30, min. dist=0.5, seed. use=10.” Finally, This study visualized the reduced-dimensional data using the Dim Plot function and displayed the distribution of feature genes within the population using the Feature Plot function.
Differential expression analysis and functional enrichment analysis: Differential expression analysis was carried out between LUAD and control samples within TCGA-LUAD dataset by ‘DESeq2’-package (v 1.38.0)12, with differentially expressed genes (DEGs) being identified based on criteria of |log2Fold Change (FC)|>0.05 and adj.p<0.05. Afterward, visually represented the DEGs associated with mitochondria and PCD through Venn diagram, and the overlapping genes were noted as candidate genes for subsequent analysis.
Candidate genes were incorporated into gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) to complete enrichment analysis via ‘cluster Profiler’-package (v 4.7.1.003)[13], to further investigate the biological functions in which these genes were involved. The cut-off criterion was set to p<0.05. Additionally, the STRING website (https://cn.string-db.org/) was utilized to probe the interaction relationships between candidate genes at the protein level (Confidence = 0.4).767.
Pseudotime developmental trajectory analysis: To infer the developmental trajectories of cell populations, utilized Monocle2 software to analyze the innate lymphoid cell (ILC) progenitor data from adult mouse bone marrow, as well as the ILC precursor data from human embryonic stages, which included ILCs from multiple human tissues during embryogenesis and ILCs from various adult tissues.
Upon selecting the appropriate datasets, the original count matrices for these populations were extracted. To construct the Cell Data Set (CDS) object from the Seurat object, three input files are required: Expression matrix information, gene information, and phenotype information. FPKM/TPM values are typically log-normally distributed, while UMI or read counts are better modeled using a negative binomial distribution.
RESULTS AND DISCUSSION
Investigating the developmental differences in the lineage of innate lymphoid progenitor cells between mice and humans: The study analyzed the development process of innate lymphoid cells (ILCs) in the adult mouse bone marrow. Through dimensionality reduction clustering and differential gene analysis, then identified five distinct populations in the adult mouse bone marrow: All-lymphoid progenitors (ALP), specified early innate lymphoid progenitor 1 (sEILP1), specified early innate lymphoid progenitor 2 (sEILP2), committed early innate lymphoid progenitor (cEILP), and innate lymphoid cell progenitors (ILCPs) (Fig. 1a). According to previous reports, in the adult bone marrow, the early innate lymphoid progenitor sEILP1 can differentiate not only into ILCPs to form the innate lymphocyte lineage but also into sEILP2, which has the potential to differentiate into dendritic cells16-20. Furthermore, differential gene expression results showed that ALP highly expresses a series of genes related to multilineage development, including Mef2c, which is essential for the normal development of megakaryocytes and platelets as well as B-cell production in the bone marrow; Dntt, which plays an important role in B-cell and T-cell maturation; and Flt3, a tyrosine-protein kinase that serves as a cell surface receptor for the cytokine FLT3LG, regulating the differentiation, proliferation, and survival of hematopoietic progenitor cells and dendritic cells (Fig. 1b)21-24. The sEILP1 highly expresses Nfil3, a gene previously reported to play a crucial role in the development of the biological clock and possesses DNA-binding transcription factor activity and transcriptional repressor activity. Tyrobp, highly expressed in sEILP2, is reported to bind non-covalently to activation receptors found on the surface of various immune cells. Its combination with TREM2 acts on monocyte-derived dendritic cells, mediating the upregulation of chemokine receptor CCR7 and promoting dendritic cell maturation and survival, further suggesting that sEILP2 has dendritic cell potential. The high expression of Tcf7 (encoding TCF-1) in cEILP is essential for the differentiation of early ILC progenitor cells25-28. It provides positive regulation for key genes in the differentiation into the ILCP lineage while inhibiting the expression of genes related to dendritic cell differentiation, thereby strengthening the cell's commitment to the ILCP lineage. These results demonstrate the typical gene expression characteristics of each cell population, indicating the accuracy of cell identity identification29.
Then the development of innate lymphocytes in the embryonic liver was analyzed. And included four embryonic liver samples taken at E11, E12, E13, and E14 (Fig. 1c). The five ILC developmental populations present in adult bone marrow also exist in the embryonic liver. Additionally, the two distinct fate specialization trajectories of ILC development were also clearly observed in the embryonic liver: One path specializing towards a DC fate (ALP>sEILP1>sEILP2) and the other towards an ILC fate (ALP-sEILP1-cEILP-ILCP) (Fig. 1d)30-32. Furthermore, analyzed the differential genes among these five precursor cell populations in the embryonic liver of mice. It was discovered that Notch1 and Ighm were highly expressed in ALP, where Notch1 signaling is involved in T-cell development, and Ighm encodes for B-lymphocyte development (Fig. 1e). Among the differentially expressed genes in sEILP1, Ctr9, a component of the PAF1 complex (PAF1C), plays multiple roles in RNA polymerase II transcription and is involved in regulating embryonic stem cell pluripotency development and maintenance33. The Ccnd2, a cell cycle-related gene, is essential for the G1/S phase transition. Additionally, Id2 in sEILP2 is associated with the differentiation of conventional type 1 dendritic cells34.
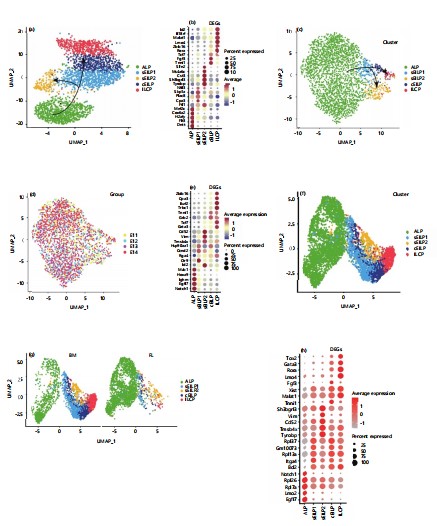
|
(a) Dots represent cells; colors indicate populations (ALP, sEILP1, sEILP2, cEILP, and ILCP), (b) Dot size: Expression proportion; color gradient (-1 to 1): Average expression, (c) Dots represent cells; colors indicate populations (ALP, sEILP1, sEILP2, cEILP, ILCP), (d) Dots represent cells; colors indicate stages (E11, E12, E13, E14), (e) Dot size: Expression proportion; color gradient (-1 to 1): Average expression, (f) Dots represent cells; colors indicate populations (ALP, sEILP1, sEILP2, cEILP, ILCP), (g) Dots represent cells; colors indicate sources and (h) Dot size: Expression proportion; color gradient (-1 to 1): Average expression
To further compare the similarities and differences between the innate lymphoid precursor cell populations in the embryonic liver and adult bone marrow, integrated the two datasets using the Harmony package in Seurat software. From the dimensionality reduction plots. This study found that the corresponding innate lymphocyte precursor populations clustered together (Fig. 1f-g). This indicates that the innate lymphoid precursor cell populations in the embryonic liver and adult bone marrow are overall similar in terms of lineage developmental trajectory35-37.
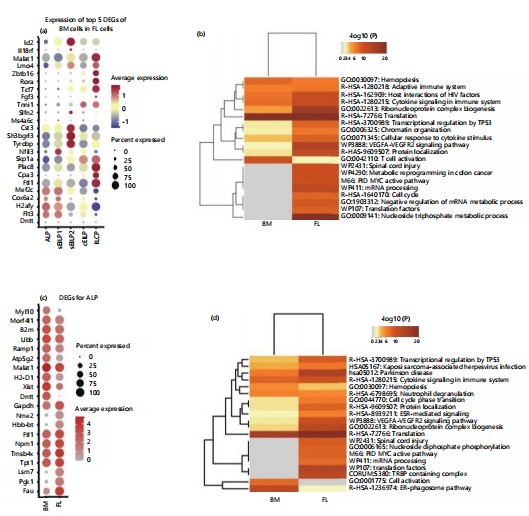
Differences between precursor populations in embryonic liver and adult bone marrow: To compare the similarities and differences in differentially expressed genes between the five precursor populations with similar developmental relationships in embryonic liver and adult bone marrow, projected the top five differentially expressed genes from each population in adult bone marrow onto the corresponding populations in the embryonic liver (Fig. 2a)38. This illustrates the expression of genes differentially expressed in adult bone marrow cell populations in their corresponding embryonic liver populations. The results showed that there is common differentially expressed genes between corresponding populations in bone marrow and embryonic liver, such as Tyrobp and Cst3, which are highly expressed in sEILP2, and Zbtb16, Rora, and Tcf7, which are highly expressed in ILCP39,40. However, there are also significant differences in the expression of differentially expressed genes between corresponding populations in bone marrow and embryonic liver. For example, genes such as Dntt and Cox6a2 are highly expressed in adult bone marrow ALP but have lower expression levels in embryonic liver ALP41-43. This suggests both differences and consistencies between progenitor cell populations in adult bone marrow and embryonic liver.
Subsequently, further analyzed the differences between the fetal liver and bone marrow ILC-related populations (Fig. 2b). The results showed that compared with adult bone marrow, the embryonic liver was enriched with functions related to mRNA processing, negative regulation of mRNA metabolism, and cell cycle. In addition, cytokine signaling function in the adaptive immune system was highly enriched in both embryonic liver and adult bone marrow44-46.
The differences between corresponding populations in different ILC developmental precursor cells in the fetal liver and bone marrow were compared. For the ALP population, differential gene expression results showed (Fig. 2c) that Fau, which is related to ribosome subunit assembly and function, is specifically overexpressed in ALP in the embryonic liver. The Lsm7, as a component of the catalytic spliceosome, plays a role in mRNA splicing. The protein encoded by Tpt1 is involved in cell growth and proliferation47. Malat1, which is overexpressed in ALP in bone marrow, can serve as a transcriptional regulator for multiple genes, including those related to cell migration. Functional enrichment analysis indicated (Fig. 2d) that ALP in the embryonic liver is enriched in functions related to mRNA processing, ribosome subunits, and pathway activation, while ALP in adult bone marrow specifically enriches functions related to cell activation.
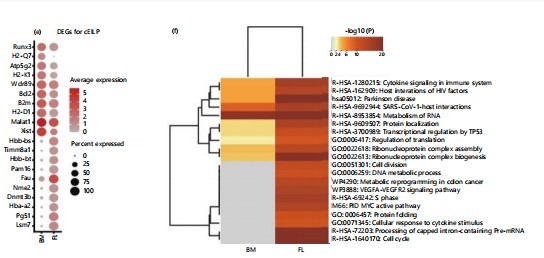
These differences are even more pronounced in cEILP, as can be seen from the differential gene expression (Fig. 2e). In the cEILP of adult bone marrow, Fau is overexpressed. This gene is associated with mucosal innate immune responses or organ- or tissue-specific immune responses. Additionally, in the cEILP of adult bone marrow, Bcl2 is overexpressed, which is related to T-cell lineage differentiation. Functional enrichment analysis reveals (Fig. 2f) that in the embryonic liver, there is an enrichment of functions related to mRNA processing, negative regulation of mRNA metabolic processes, and cell cycle48-50.
Overall, the development of ILCs in the embryonic liver and adult bone marrow exhibits a high degree of consistency, with the same developmental hierarchy. However, there are some differences, mainly reflected in the higher expression of genes related to cell cycle, cell proliferation, and mRNA processing in the ILC-related populations of the embryonic liver compared to those of the adult bone marrow51. This suggests that compared to bone marrow, the lymphoid progenitor cell populations and ILCP precursor cells in the fetal liver are in a more proliferative and biosynthetically active cellular state.
|
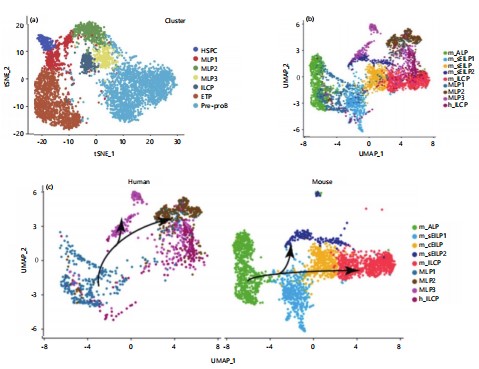
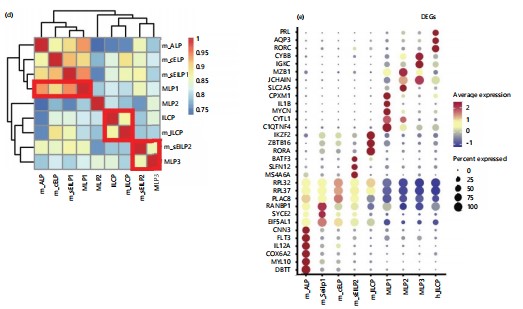
Inter-species comparison of innate lymphocyte development between humans and mice neutrophil heterogeneity: After analyzing the developmental process of ILCs in both embryonic and adult stages of mice, proceed to explore the conservation of ILC specialization between humans and mice and the similarities and differences that exist. Next, further compared the inter-species similarities and differences in ILC development between humans and mice. Firstly, a deeper analysis of data in the laboratory in the early stage was conducted, elucidating the specialization pathway of ILCs or DCs in the fetal liver, specifically MLP1>MLP2>ILCP/MLP3. From the results, it can be observed that HSPCs are located at the starting point of differentiation, with MLP1 positioned downstream of HSPCs51-53. The MLP1 exhibits a bifurcation in fate differentiation: One path leads towards MLP2 and MLP3 (pointing towards the B-cell and ILCP directions), while the other leads towards the T-cell direction (Fig. 3a). This studyfurther conducted an integrated analysis of human and mouse data. Through CCA integration of data from different sources. This studyobserved clustering of mouse ALP and human MLP1, as well as mouse sEILP2 and human MLP3, in the population dimensionality reduction plot (Fig. 3b). This suggests a correspondence in the developmental process between human embryonic MLP1 and mouse ALP populations and a possible correspondence between human embryonic MLP3 and mouse sEILP2. Furthermore, distinguished between mouse and human sources to examine the dimensionality reduction results among populations separately (Fig. 3c). From the figure, distinct fate specialization trajectories can be observed for both humans and mice). In humans, MLP1 develops into MLP2, which further specializes in MLP3 or ILCP. In mice, ALP develops into sEILP1, which then further differentiates into sEILP2 or specializes into ILCP via cEILP54. The study further examined the tissue distribution of all populations, and the tissue information was reasonably distributed among the populations. More naive progenitor cell populations mainly originated from bone marrow and fetal liver, indicating no batch effects due to tissue site.
This also conducted a correlation analysis of ILCP-related precursor cell populations between humans and mice (Fig. 3d). Consistent with the results of the integrated analysis, This study found that the correspondence between human and mouse ILCP development-related populations is as follows: MLP1 in humans corresponds to ALP and sEILP1 in mice, all of which commonly express characteristic genes of lymphoid progenitor cells; MLP3 in humans corresponds to sEILP2 in mice (Fig. 3f)55-59. Interestingly, MLP2 in humans and cEILP in mice are located upstream of human and mouse ILCPs, respectively, but they do not have corresponding populations across species. This reflects differences in the developmental process of the ILCP lineage between humans and mice.
Analyze the conservativeness and differences in gene expression patterns during the fate selection process of human and mouse ILCs: This study further explored the conservativeness and differences in gene expression patterns during the fate selection process of human and mouse ILCs. The expression changes of key transcription factors for ILC development in humans and mice along the developmental sequences were studied (Fig. 4a). The results showed that transcription factors such as Irf8, Spi1, Mef2c, Lyl1, and Flt3 were progressively downregulated during the specialization of ILCs in mice, while factors such as Runx3, Tcf7, Tox2, and Id2 were progressively upregulated along the developmental trajectory (Fig. 4b)60-62.
During the specialization of human ILC fate, the expression of transcription factors SPI1, LMO2, BCL11A, and LYL1 shows a gradual downregulation, while factors such as ID2, TOX2, TCF7, and LMO4 exhibit a progressive upregulation along the developmental sequence (Fig. 4c). In summary, during the specialization of ILCs, myeloid differentiation-related transcription factors such as Lmo2, Spi1, and Lyl1 are downregulated in both humans and mice during the specialization of ILC precursor cell populations. Meanwhile, key transcription factors for ILC development, including Id2, Tox2, and Tcf7, are upregulated in both species, indicating the conservativeness of transcriptional regulation during the specialization of human and mouse ILCs. On the other hand, there are also species differences in the development of ILCs between humans and mice63,64. For instance, the expression of Bcl11a and Tcf4 gradually decreases in mice but shows no significant change in humans. Additionally, Zbtb16 and Rora are upregulated during mouse ILC specialization, but no upregulation trend is observed in humans. This suggests differences in early ILC development between humans and mice.
|
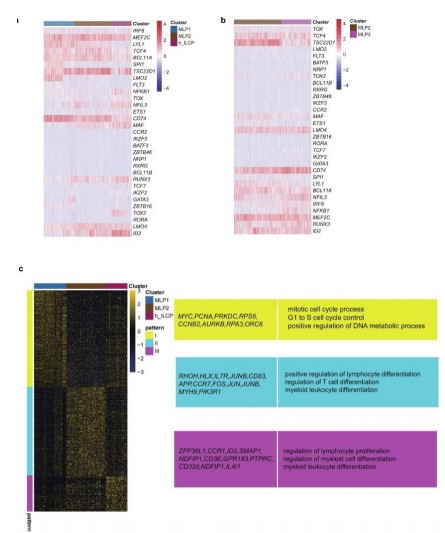
The current study further explores the changes in gene expression patterns during the lineage development of human and mouse innate lymphoid cell (ILC) progenitor cells. In mice, genes with dynamically changing expression levels during the differentiation process from the all-lymphoid progenitors (ALPs) to innate lymphoid cell progenitors (ILCPs) can be categorized into four patterns. Then present typical functions related to the developmental process for each pattern. The ALP population corresponds to Pattern 1, where functions such as immune response involving B cells, immune reactions involving lymphocyte activation, and positive regulation of T-cell proliferation are predominantly enriched. The sEILP1 population corresponds to Pattern 2, mainly enriching functions like cell cycle, DNA replication, and messenger RNA processing. The cEILP population corresponds to Pattern 3, primarily enriching functions such as lymphocyte activation, leukocyte activation, and negative regulation of cell proliferation. The ILCP population corresponds to Pattern 4, predominantly enriching functions including leukocyte activation and regulation of lymphocyte differentiation (Fig. 4c).
During the differentiation process from multipotent lymphoid progenitors 1 (MLP1) to innate lymphoid cell progenitors (ILCPs) in humans, dynamically changing gene expression levels can be categorized into three patterns. Pattern 1, corresponding to MLP1, predominantly enriches functions such as cell cycle processes, cell cycle regulation, and positive regulation of DNA metabolic processes. Pattern 2, associated with MLP2, mainly focuses on functions like positive regulation of lymphocyte differentiation, regulation of T-cell differentiation, and myeloid leukocyte differentiation. Pattern 3, linked to ILCPs, primarily involves the regulation of lymphocyte differentiation and myeloid cell differentiation (Fig. 4d). In summary, although there is a correspondence between the all-lymphoid progenitors (ALP) population in mice and the MLP1 population in humans, there are differences in terms of differentially expressed genes. Functional enrichment results indicate that the ALP population mainly enriches functions related to the proliferation and differentiation of white blood cells such as B-cells, T-cells, and lymphocytes, while MLP1 predominantly exhibits characteristics related to cell cycle and cell proliferation65. The sEILP1 population mainly focuses on functions such as cell cycle, DNA replication, and messenger RNA processing. This functionally suggests differences in the specialization process of ILCs between humans and mice.
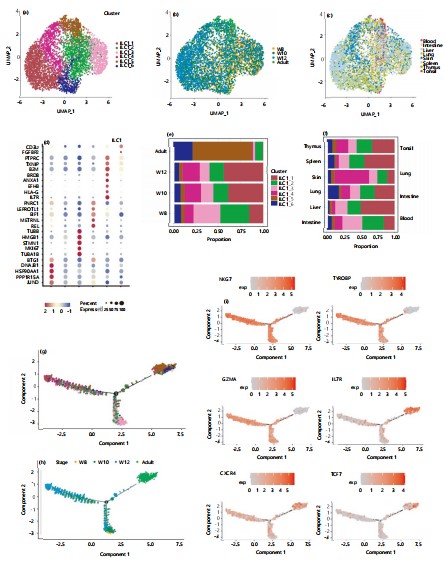
Heterogeneity of group 1 innate lymphoid cells during embryonic development and comparison with adulthood: After analyzing the development of group 1 innate lymphoid cells (ILC1s) during embryonic stages, given the crucial role ILC1s play in this period, This study sought to investigate the heterogeneity of ILC1s across different tissues during embryonic development and to compare the differences between ILC1s in embryonic and adult stages. To achieve this, This studied single-cell sequencing data of innate lymphoid cells from various tissues in both human and mouse embryos as well as adult stages. Group 1 innate lymphoid cells can be further subdivided into six distinct subpopulations: ILC1_1, ILC1_2, ILC1_3, ILC1_4, ILC1_5, and ILC1_6 (Fig. 5a). The distribution across tissues and developmental time points for these subpopulations is illustrated in Fig. 5b-c. From the time-point distribution of group 1 innate lymphoid cells, ILC1_5 is predominantly found in adult stages. Comparative analysis of each subpopulation revealed that ILC1_5 specifically expresses genes characteristic of more mature cells, such as HLA-G and IL7R (Fig. 5d). The data also show that both ILC1_2 and ILC1_3 subpopulations exhibit a decreasing trend in cell numbers over time (Fig. 5e). Notably, ILC1_3 highly expresses genes involved in the regulation of DNA replication, including TUBA1B, TUBB, TOP2A, and MKI67 (Fig. 5d), suggesting that ILC1_3 may represent a more immature cell population. The ILC1_4 shows an increasing trend in cell numbers during embryonic development, while ILC1_6 expands in adulthood and specifically expresses FGFBP2, a hallmark gene of natural killer (NK) cells. This suggests that ILC1_6 may include cells that are difficult to distinguish from NK cells within group 1 innate lymphoid cells. Interestingly, current data analysis reveals tissue-specific distribution biases among the subpopulations of group 1 innate lymphoid cells (ILC1s) during embryonic development. Notably, ILC1_4 primarily originates from embryonic skin tissue, while ILC1_6 is predominantly derived from blood and primarily found in the adult stage. This study hypothesizes that ILC1_6 represents a more mature state, gradually specialized from blood tissue. The ILC1_5 consists mostly of adult cells, with the highest proportion found in the tonsils (Fig. 5f). The study simulated the developmental trajectory of group 1 innate lymphoid cells using Monocle 2. Consistent with our expectations, the inferred pseudo-time trajectory begins with ILC1_3 (the presumed immature subpopulation) and subsequently bifurcates into two developmental paths. One path differentiates into a branch characterized by ILC1_1, representing group 1 innate lymphoid cells with a broad tissue distribution during embryonic stages. The other path diverges into a branch dominated by ILC1_5 and ILC1_6, exhibiting adult-stage characteristics (Fig. 5g). In terms of projected time, the trajectory shows that differentiation begins around the seventh and eighth weeks, with one branch diverging toward the later embryonic stage and the other toward adulthood. The fitted developmental trajectory aligns well with known biological processes (Fig. 5h). By highlighting transcription factors differentially expressed between the embryonic and adult stages, This study found that NKG7, TYROBP, and GZMA which are embryonic stage-specific are highly expressed in the ILC1_1-dominated branch. Conversely, adult stage-specific transcription factors such as IL7R, CXCR4, and TCF7 are highly expressed in the ILC1_5 and ILC1_6-dominated branches. This further validates that our pseudo-time axis accurately reflects the biological processes (Fig. 5i).
|
|
Innate lymphoid cells (ILCs) have garnered significant attention in recent decades as a crucial component of innate immunity, primarily residing in mucosal tissues such as the gut, lungs, and skin. The ILCs can be categorized into three distinct groups based on their function, secreted cytokines, and the transcription factors required for differentiation: Group 1 ILCs (including natural killer cells and ILC1), ILC2, and Group 3 ILCs (comprising ILC3 and lymphoid tissue inducer cells). In recent years, with the deepening understanding of ILCs, there has been increasing emphasis on their immunological roles within the immune system. Studies in mice have demonstrated the significant role of ILCs in chronic inflammation, infection, allergies, and cancer. Recent research also indicates their functions in maintaining tissue homeostasis, promoting tissue repair, and regulating tissue inflammation. However, the development and origin of ILCs remain partly unknown. For instance, while the ILC lineage development in the bone marrow of adult mice is relatively well understood, the origin and developmental pathway of ILCs during the embryonic stage of mice are still unclear. This study first analyzed the developmental process and fate specialization trajectory of ILCs in the fetal liver and then compared the similarities and differences between adult and embryonic ILC lineage development in mice. Furthermore, explored the differences between humans and mice, revealing unique regulatory mechanisms of ILC development in humans.
In the bone marrow of adult mice, the ILC lineage developmental pathway begins with all-lymphoid progenitors (ALPs), which then develop into sEILP1. The sEILP1 has a fate specialization branch, where one path can differentiate into sEILP2 (with dendritic cell potential), and the other path differentiates into cEILP, which further differentiates into ILCP. By analyzing published data on mouse fetal livers, This study discovered the presence of five ILC developmental populations: ALP, sEILP1, sEILP2, cEILP, and ILCP. The fetal liver of mice exhibits similar ILC fate specialization and selection pathways as the bone marrow: One path specializes towards a DC fate, while the other specializes towards an ILCP fate. Additionally, This study found that fetal liver-derived populations express higher levels of genes related to cell cycle, cell proliferation, and mRNA processing compared to bone marrow populations in adults, suggesting differences in ILC fate differentiation between the fetal liver and bone marrow.
This study further compared the similarities and differences in ILC specialization between humans and mice. Firstly, revealed the corresponding relationships between human and mouse ILC developmental populations. Integrated analysis and correlation analysis showed that ALP and sEILP1 in mice correspond to MLP1 in humans, sEILP2 in mice corresponds to MLP3 in humans, and ILCP in humans corresponds to ILCP in mice, indicating conservation in ILC development between the two species. However, MLP2 in humans, as the precursor cell of ILCP, does not have a clear corresponding relationship with cEILP, the precursor cell of ILCP in mice, suggesting differences in ILC development between humans and mice. These results indicate both conservation and divergence in the lineage hierarchy of ILC development between humans and mice. Furthermore, there are also species-specific conservation and divergence in gene expression during ILC development. For example, both ALP and MLP1 highly express genes associated with lymphoid progenitor cells. However, ALP highly expresses a series of genes related to multilineage development, including Mef2c (megakaryocytes and B-cells), Dntt (T and B-cell development), and Flt3 (lymphoid progenitor cells and dendritic cells), which are not expressed in MLP1. Additionally, this study compared changes in typical transcription factors during ILC lineage development. The results showed significant differences in transcription factor changes between humans and mice. In humans, factors such as IFR8 and TCF4 gradually decrease along the developmental sequence, while factors like ID2, NFIL3, and MAF gradually increase. In mice, factors like Irf8 and Spi1 decrease progressively along the developmental trajectory, while factors such as Tcf7 and Tox2 increase progressively. These findings highlight the species-specific conservation and divergence at the cellular and molecular levels during ILC development in humans and mice.
CONCLUSION
This study, elucidated the developmental trajectory and fate specialization of ILCs in mouse fetal liver, revealing similarities and differences compared to adult bone marrow. The study identified conserved and divergent features of ILC development between humans and mice, including lineage hierarchy, gene expression patterns, and transcription factor dynamics. Specifically, it demonstrated that fetal liver ILC populations exhibit enhanced cell cycle and proliferation-related gene expression compared to adult bone marrow. Furthermore, the study uncovered species-specific conservation and divergence in ILC lineage development, highlighting unique regulatory mechanisms in humans. These findings provide new insights into the developmental origins and evolutionary adaptations of ILCs, offering a foundation for further exploration of their roles in immunity and disease.
SIGNIFICANCE STATEMENT
Innate lymphoid cells (ILCs) play a vital role in immune homeostasis and tissue repair, but their developmental pathways, especially in humans, are not fully understood. This study maps the development of ILCs in the mouse fetal liver, revealing key insights into the differentiation processes and their comparison with human ILC development. By identifying species-specific differences and similarities, the research uncovers critical molecular regulators and provides a more comprehensive understanding of ILC specialization. These findings not only advance our knowledge of immune cell development but also open the door to better strategies for in vitro ILC regeneration and understanding tissue-specific immune responses, with potential implications for immune-related diseases and therapeutic interventions.
REFERENCES
- Vivier, E., D. Artis, M. Colonna, A. Diefenbach and J.P.D. Santo et al., 2018. Innate lymphoid cells: 10 years on. Cell, 174: 1054-1066.
- Harly, C., D. Kenney, G. Ren, B. Lai and T. Raabe et al., 2019. The transcription factor TCF-1 enforces commitment to the innate lymphoid cell lineage. Nat. Immunol., 20: 1150-1160.
- Poli, A., T. Michel, M. Thérésine, E. Andrès, F. Hentges and J. Zimmer, 2009. CD56bright natural killer (NK) cells: An important NK cell subset. Immunology, 126: 458-465.
- Montaldo, E., G.D. Zotto, M.D. Chiesa, M.C. Mingari, A. Moretta, A. de Maria and L. Moretta, 2013. Human NK cell receptors/markers: A tool to analyze NK cell development, subsets and function. Cytometry Part A, 83A: 702-713.
- Tang, Y., C. Peitzsch, H.N. Charoudeh, M. Cheng and P. Chaves et al., 2012. Emergence of NK-cell progenitors and functionally competent NK-cell lineage subsets in the early mouse embryo. Blood, 120: 63-75.
- Bai, L., M. Vienne, L. Tang, Y. Kerdiles and M. Etiennot et al., 2021. Liver type 1 innate lymphoid cells develop locally via an interferon-γ-dependent loop. Science, 371.
- Chen, Y., X. Wang, X. Hao, B. Li and W. Tao et al., 2022. Ly49E separates liver ILC1s into embryo-derived and postnatal subsets with different functions. J. Exp. Med., 219.
- Sparano, C., D. Solís-Sayago, A. Vijaykumar, C. Rickenbach and M. Vermeer et al., 2022. Embryonic and neonatal waves generate distinct populations of hepatic ILC1s. Sci. Immunol., 7.
- Constantinides, M.G., H. Gudjonson, B.D. McDonald, I.E. Ishizuka, P.A. Verhoef, A.R. Dinner and A. Bendelac, 2015. PLZF expression maps the early stages of ILC1 lineage development. Proc. Natl. Acad. Sci. U.S.A., 112: 5123-5128.
- Ferreira, A.C.F., A.C.H. Szeto, M.W.D. Heycock, P.A. Clark and J.A. Walker et al., 2021. RORα is a critical checkpoint for T cell and ILC2 commitment in the embryonic thymus. Nat. Immunol., 22: 166-178.
- Kernfeld, E.M., R.M.J. Genga, K. Neherin, M.E. Magaletta, P. Xu and R. Maehr, 2018. A single-cell transcriptomic atlas of thymus organogenesis resolves cell types and developmental maturation. Immunity, 48: 1258-1270.e6.
- Stehle, C., T. Rückert, R. Fiancette, D.W. Gajdasik and C. Willis et al., 2021. T-bet and RORα control lymph node formation by regulating embryonic innate lymphoid cell differentiation. Nat. Immunol., 22: 1231-1244.
- Sánchez, M.J., M.O. Muench, M.G. Roncarolo, L.L. Lanier and J.H. Phillips, 1994. Identification of a common T/natural killer cell progenitor in human fetal thymus. J. Exp. Med., 180: 569-576.
- Renoux, V.M., A. Zriwil, C. Peitzsch, J. Michaëlsson, D. Friberg, S. Soneji and E. Sitnicka, 2015. Identification of a human natural killer cell lineage-restricted progenitor in fetal and adult tissues. Immunity, 43: 394-407.
- Liu, C., Y. Gong, H. Zhang, H. Yang and Y. Zeng et al., 2021. Delineating spatiotemporal and hierarchical development of human fetal innate lymphoid cells. Cell Res., 31: 1106-1122.
- Collins, A., N. Rothman, K. Liu and S.L. Reiner, 2017. Eomesodermin and T-bet mark developmentally distinct human natural killer cells. JCI Insight, 2.
- Arora, N., Y. Sadovsky, T.S. Dermody and C.B. Coyne, 2017. Microbial vertical transmission during human pregnancy. Cell Host Microbe, 21: 561-567.
- Vaaben, A.V., J. Levan, C.B.T. Nguyen, P.C. Callaway and M. Prahl et al., 2022. In utero activation of natural killer cells in congenital cytomegalovirus infection. J. Infect. Dis., 226: 566-575.
- Moro, K., T. Yamada, M. Tanabe, T. Takeuchi and T. Ikawa et al., 2010. Innate production of TH2 cytokines by adipose tissue-associated c-Kit+Sca-1+ lymphoid cells. Nature, 463: 540-544.
- Monticelli, L.A., G.F. Sonnenberg, M.C. Abt, T. Alenghat and C.G.K. Ziegler et al., 2011. Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nat. Immunol., 12: 1045-1054.
- Schneider, C., J. Lee, S. Koga, R.R. Ricardo-Gonzalez and J.C. Nussbaum et al., 2019. Tissue-resident group 2 innate lymphoid cells differentiate by layered ontogeny and in situ perinatal priming. Immunity, 50: 1425-1438.E5.
- Koga, S., K. Hozumi, K.I. Hirano, M. Yazawa and T. Terooatea et al., 2018. Peripheral PDGFRα+gp38+ mesenchymal cells support the differentiation of fetal liver-derived ILC2. J. Exp. Med., 215: 1609-1626.
- Bal, S.M., J.H. Bernink, M. Nagasawa, J. Groot and M.M. Shikhagaie et al., 2016. IL-1β, IL-4 and IL-12 control the fate of group 2 innate lymphoid cells in human airway inflammation in the lungs. Nat. Immunol., 17: 636-645.
- Mjösberg, J.M., S. Trifari, N.K. Crellin, C.P. Peters and C.M. van Drunen et al., 2011. Human IL-25- and IL-33-responsive type 2 innate lymphoid cells are defined by expression of CRTH2 and CD161. Nat. Immunol., 12: 1055-1062.
- Saluzzo, S., A.D. Gorki, B.M.J. Rana, R. Martins and S. Scanlon et al., 2017. First-breath-induced type 2 pathways shape the lung immune environment. Cell Rep., 18: 1893-1905.
- de Kleer, I.M., M. Kool, M.J.W. de Bruijn, M. Willart and J. van Moorleghem et al., 2016. Perinatal activation of the interleukin-33 pathway promotes type 2 immunity in the developing lung. Immunity, 45: 1285-1298.
- Fonseca, W., N.W. Lukacs, S. Elesela and C.A. Malinczak, 2021. Role of ILC2 in viral-induced lung pathogenesis. Front. Immunol., 12.
- Satoh-Takayama, N., 2016. Heterogeneity and diversity of group 3 innate lymphoid cells: New cells on the block. Int. Immunol., 28: 29-34.
- Melo-Gonzalez, F. and M.R. Hepworth, 2017. Functional and phenotypic heterogeneity of group 3 innate lymphoid cells. Immunology, 150: 265-275.
- Magri, G., M. Miyajima, S. Bascones, A. Mortha and I. Puga et al., 2014. Innate lymphoid cells integrate stromal and immunological signals to enhance antibody production by splenic marginal zone B cells. Nat. Immunol., 15: 354-364.
- Eberl, G., M. Colonna, J.P.D. Santo and A.N.J. McKenzie, 2015. Innate lymphoid cells: A new paradigm in immunology. Science, 348.
- van de Pavert, S.A., 2021. Lymphoid Tissue inducer (LTi) cell ontogeny and functioning in embryo and adult. Biomed. J., 44: 123-132.
- Mortha, A., A. Chudnovskiy, D. Hashimoto, M. Bogunovic, S.P. Spencer, Y. Belkaid and M. Merad, 2014. Microbiota-dependent crosstalk between macrophages and ILC3 promotes intestinal homeostasis. Science, 343.
- Klose, C.S.N., E.A. Kiss, V. Schwierzeck, K. Ebert and T. Hoyler et al., 2013. A T-bet gradient controls the fate and function of CCR6-RORγt+ innate lymphoid cells. Nature, 494: 261-265.
- Guo, X., J. Qiu, T. Tu, X. Yang and L. Deng et al., 2014. Induction of innate lymphoid cell-derived interleukin-22 by the transcription factor STAT3 mediates protection against intestinal infection. Immunity, 40: 25-39.
- Hepworth, M.R., L.A. Monticelli, T.C. Fung, C.G.K. Ziegler and S. Grunberg et al., 2013. Innate lymphoid cells regulate CD4+ T-cell responses to intestinal commensal bacteria. Nature, 498: 113-117.
- Pearson, C., E.E. Thornton, B. McKenzie, A.L. Schaupp and N. Huskens et al., 2016. ILC3 GM-CSF production and mobilisation orchestrate acute intestinal inflammation. eLife, 5.
- Zeng, B., S. Shi, G. Ashworth, C. Dong, J. Liu and F. Xing, 2019. ILC3 function as a double-edged sword in inflammatory bowel diseases. Cell Death Dis., 10.
- Eberl, G., S. Marmon, M.J. Sunshine, P.D. Rennert, Y. Choi and D.R. Littman, 2004. An essential function for the nuclear receptor RORγt in the generation of fetal lymphoid tissue inducer cells. Nat. Immunol., 5: 64-73.
- Eberl, G., 2012. Development and evolution of RORγt+ cells in a microbe’s world. Immunol. Rev., 245: 177-188.
- Scandella, E., B. Bolinger, E. Lattmann, S. Miller and S. Favre et al., 2008. Restoration of lymphoid organ integrity through the interaction of lymphoid tissue-inducer cells with stroma of the T cell zone. Nat. Immunol., 9: 667-675.
- Ansel, K.M., V.N. Ngo, P.L. Hyman, S.A. Luther and R. Förster et al., 2000. A chemokine-driven positive feedback loop organizes lymphoid follicles. Nature, 406: 309-314.
- Simic, M., I. Manosalva, L. Spinelli, R. Gentek and R.R. Shayan et al., 2020. Distinct waves from the hemogenic endothelium give rise to layered lymphoid tissue inducer cell ontogeny. Cell Rep., 32.
- Cherrier, M., S. Sawa and G. Eberl, 2012. Notch, Id2, and RORγt sequentially orchestrate the fetal development of lymphoid tissue inducer cells. J. Exp. Med., 209: 729-740.
- Possot, C., S. Schmutz, S. Chea, L. Boucontet, A. Louise, A. Cumano and R. Golub, 2011. Notch signaling is necessary for adult, but not fetal, development of RORγt+ innate lymphoid cells. Nat. Immunol., 12: 949-958.
- van de Pavert, S.A., M. Ferreira, R.G. Domingues, H. Ribeiro and R. Molenaar et al., 2014. Maternal retinoids control type 3 innate lymphoid cells and set the offspring immunity. Nature, 508: 123-127.
- van de Pavert, S.A., B.J. Olivier, G. Goverse, M.F. Vondenhoff and M. Greuter et al., 2009. Chemokine CXCL13 is essential for lymph node initiation and is induced by retinoic acid and neuronal stimulation. Nat. Immunol., 10: 1193-1199.
- de Agüero, M.G., S.C. Ganal-Vonarburg, T. Fuhrer, S. Rupp and Y. Uchimura et al., 2016. The maternal microbiota drives early postnatal innate immune development. Science, 351: 1296-1302.
- Qiu, J., J.J. Heller, X. Guo, Z.M.E. Chen, K. Fish, Y.X. Fu and L. Zhou, 2012. The aryl hydrocarbon receptor regulates gut immunity through modulation of innate lymphoid cells. Immunity, 36: 92-104.
- Meier, D., C. Bornmann, S. Chappaz, S. Schmutz and L.A. Otten et al., 2007. Ectopic lymphoid-organ development occurs through interleukin 7-mediated enhanced survival of lymphoid-tissue-inducer cells. Immunity, 26: 643-654.
- Jones, R., E.J. Cosway, C. Willis, A.J. White and W.E. Jenkinson et al., 2018. Dynamic changes in intrathymic ILC populations during murine neonatal development. Eur. J. Immunol., 48: 1481-1491.
- Marquardt, N., M.A. Ivarsson, E. Sundström, E. Åkesson and E. Martini et al., 2016. Fetal CD103+ IL-17-producing group 3 innate lymphoid cells represent the dominant lymphocyte subset in human amniotic fluid. J. Immunol., 197: 3069-3075.
- Forkel, M., L. Berglin, E. Kekäläinen, A. Carlsson and E. Svedin et al., 2017. Composition and functionality of the intrahepatic innate lymphoid cell-compartment in human nonfibrotic and fibrotic livers. Eur. J. Immunol., 47: 1280-1294.
- Cupedo, T., N.K. Crellin, N. Papazian, E.J. Rombouts and K. Weijer et al., 2009. Human fetal lymphoid tissue-inducer cells are interleukin 17-producing precursors to RORC+ CD127+ natural killer-like cells. Nat. Immunol., 10: 66-74.
- Robinette, M.L., A. Fuchs, V.S. Cortez, J.S. Lee and Y. Wang et al., 2015. Transcriptional programs define molecular characteristics of innate lymphoid cell classes and subsets. Nat. Immunol., 16: 306-317.
- Shikhagaie, M.M., Å.K. Björklund, J. Mjösberg, J.S. Erjefält and A.S. Cornelissen et al., 2017. Neuropilin-1 is expressed on lymphoid tissue residing LTi-like group 3 innate lymphoid cells and associated with ectopic lymphoid aggregates. Cell Rep., 18: 1761-1773.
- Gray, J., K. Oehrle, G. Worthen, T. Alenghat, J. Whitsett and H. Deshmukh, 2017. Intestinal commensal bacteria mediate lung mucosal immunity and promote resistance of newborn mice to infection. Sci. Transl. Med., 9. https://doi.org/10.1126/scitranslmed.aaf9412
- Oherle, K., E. Acker, M. Bonfield, T. Wang and J. Gray et al., 2020. Insulin-like growth factor 1 supports a pulmonary niche that promotes type 3 innate lymphoid cell development in newborn lungs. Immunity, 52: 275-294.E9.
- Hernández-Torres, D.C. and C. Stehle, 2022. Embryonic ILC-poiesis across tissues. Front. Immunol., 13.
- Yudanin, N.A., F. Schmitz, A.L. Flamar, J.J.C. Thome and E.T. Wojno et al., 2019. Spatial and temporal mapping of human innate lymphoid cells reveals elements of tissue specificity. Immunity, 50: 505-519.E4.
- Colonna, M., 2018. Innate lymphoid cells: Diversity, plasticity, and unique functions in immunity. Immunity, 48: 1104-1117.
- Ding, Y. and A. Bhandoola, 2021. Building early defenses. Cell Res., 31: 1041-1042.
- Harly, C., M. Cam, J. Kaye and A. Bhandoola, 2018. Development and differentiation of early innate lymphoid progenitors. J. Exp. Med., 215: 249-262.
- Yang, Q., F. Li, C. Harly, S. Xing and L. Ye et al., 2015. TCF-1 upregulation identifies early innate lymphoid progenitors in the bone marrow. Nat. Immunol., 16: 1044-1050.
- Heinz, S., C. Benner, N. Spann, E. Bertolino and Y.C. Lin et al., 2010. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell, 38: 576-589.
How to Cite this paper?
APA-7 Style
Yahui,
H., Yujing,
Y., Guoli,
Y., Yuefeng,
Z. (2025). Comparative Single-Cell Transcriptome Analysis of Early Innate Lymphoid Cell Development in Humans and Mice. Scientific Asia, 1(1), 16-32. https://doi.org/10.3923/sa.2025.16.32
ACS Style
Yahui,
H.; Yujing,
Y.; Guoli,
Y.; Yuefeng,
Z. Comparative Single-Cell Transcriptome Analysis of Early Innate Lymphoid Cell Development in Humans and Mice. Scientific Asia 2025, 1, 16-32. https://doi.org/10.3923/sa.2025.16.32
AMA Style
Yahui
H, Yujing
Y, Guoli
Y, Yuefeng
Z. Comparative Single-Cell Transcriptome Analysis of Early Innate Lymphoid Cell Development in Humans and Mice. Scientific Asia. 2025; 1(1): 16-32. https://doi.org/10.3923/sa.2025.16.32
Chicago/Turabian Style
Yahui, Han, Yang Yujing, Yao Guoli, and Zhang Yuefeng.
2025. "Comparative Single-Cell Transcriptome Analysis of Early Innate Lymphoid Cell Development in Humans and Mice" Scientific Asia 1, no. 1: 16-32. https://doi.org/10.3923/sa.2025.16.32

This work is licensed under a Creative Commons Attribution 4.0 International License.